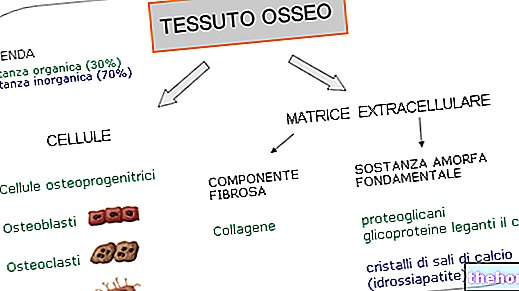
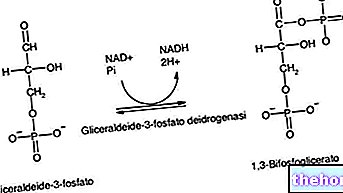
De synthese van vetzuren begint bij acetyl-co-enzym A en komt ruwweg overeen met het omgekeerde pad van hun afbraak; bij de synthese van vetzuren wordt een reeks bicarbonaatfragmenten toegevoegd aan het uitgangsacetyl-co-enzym A.
De synthese van vetzuren is volledig cytoplasmatisch (dwz de enzymen die deze synthese katalyseren worden gevonden in het cytoplasma). Het acetyl-co-enzym A dat in het cytoplasma wordt gebruikt voor de synthese van vetzuren is van mitochondriale oorsprong: een klein deel wordt getransporteerd door carnitine, door de werking van twee acyltransferase-enzymen (een cytoplasmatisch en een mitochondriaal) en een translocase-enzym. co-enzym A van mitochondriale oorsprong wordt verkregen via een gespecialiseerde route: de citraatlyase (de naam is afgeleid van het eerste enzym van dit pad).
Acetylco-enzym A aanwezig in mitochondriën is afkomstig van glycolyse na de werking van pyruvaatdehydrogenase; Acetyl-co-enzym A ondergaat de werking van het enzym citraatsynthase: dit enzym katalyseert de vorming van citraat door reactie van acetyl-co-enzym A met oxaalacetaat.Als de krebs-cyclus in staat is om aan de energiebehoeften te voldoen, kan een deel van het citraat (de hoeveelheid die niet nodig is voor de krebs-cyclus) de mitochondriën verlaten en het cytoplasma bereiken, waar het citraatlyase-enzym, dat energie verbruikt, het weer omzet in acetyl-co-enzym A en oxaalacetaat Op deze manier is het mogelijk om acetyl co-enzym A beschikbaar te hebben in het cytoplasma, maar het gevormde oxaalacetaat moet terug naar de mitochondriën om weer beschikbaar te zijn voor het citraatsynthase-enzym.
Het oxaalacetaat wordt vervolgens omgezet in malaat door de werking van het enzym malaatdehydrogenase cytoplasmatisch (een cytoplasmatisch NADH wordt verbruikt): het malaat is een permeabele metaboliet en kan opnieuw de mitochondriën binnendringen waar het, onder de werking van het mitochondriale malaatdehydrogenase-enzym, opnieuw wordt omgezet in oxaalacetaat (er wordt ook een NADH verkregen); de cytoplasmatische patiënt kan als alternatief de werking ondergaan van het appelzuurenzym, dat een decarboxylering en dehydrogenering uitvoert, om te worden omgezet in pyruvaat. Het appelzuur-enzym werkt op NADP + (het is vergelijkbaar met nicotinamide-adeninedinucleotide, maar in tegenstelling tot dit heeft het een fosforgroep op de tweede hydroxylgroep op een van de twee ribose-eenheden), daarom wordt in de overgang van malaat naar pyruvaat NADPH geproduceerd ( dat wordt gebruikt bij de biosynthese) Pyruvaat komt vervolgens de mitochondriën binnen waar het wordt omgezet in oxaalacetaat door de werking van pyruvaatcarboxylase of in acetyl-co-enzym A via pyruvaatdehydrogenase.
Laten we een voorbeeld bekijken: acht moleculen acetyl-co-enzym A zijn nodig om palmitinezuur (keten met zestien koolstofatomen) te synthetiseren, maar slechts één ervan wordt als zodanig gebruikt: zeven moleculen acetyl-co-enzym A worden omgezet in malonyl-co-enzym A door het " enzym acetyl-co-enzym A-carboxylase (dit enzym gebruikt een CO2-molecuul en heeft biotine als cofactor).
Het acetyl-co-enzym Een carboxylase-enzym kan voorkomen in een bijna inactieve gedispergeerde vorm en een actieve aggregaatvorm (ongeveer twintig eenheden); de overgang van de gedispergeerde naar de aggregaatvorm vindt plaats wanneer er in het cytoplasma een "hoge concentratie citraat is: citraat wordt een positieve modulator van het acetyl-co-enzym A-carboxylase-enzym.
Het acetyl-co-enzym A-carboxylase-enzym heeft andere positieve (insuline) en negatieve (glucagon, adrenaline en acyl-co-enzym A) modulatoren.
We zullen de synthese van vetzuren in de bacterie escherichia coli analyseren, waarbij deze synthese plaatsvindt door de werking van zeven verschillende eiwitten; in eukaryote cellen is het mechanisme waarmee de synthese van vetzuren plaatsvindt vergelijkbaar met dat van bacteriën, maar in eukaryoten zijn de zeven enzymen die verantwoordelijk zijn voor de synthese gegroepeerd in twee multi-enzymcomplexen A en B.
In bacteriën coderen zeven verschillende genen voor:
- ACP (acyldragereiwit);
- ACP-acetyltransacetylase;
- ACP.malonyltransacetylase;
- a-keto-acyl-ACP-synthase (condenserend enzym);
- a-keto-acyl-ACP-reductase;
- D-a-hydroxy-acyldehydratase;
- enoil-ACP geredigeerd.
In eukaryoten coderen twee genen voor:
Subeenheid A
ACS;
condenserend enzym
β-keto-acyl-ACP-reductase.
Subeenheid B
ACP-acetyltransacetylase;
ACP-malonyltransacetylase;
D-a-hydroxy-acyldehydratase;
enoil-ACP geredigeerd.
De zeven eiwitten van Escherichia coli zijn zo gerangschikt dat er een centrale is (de ACP) en de andere zes daaromheen.
Twee sulfhydrylgroepen zijn betrokken bij zijn enzymatische werking: één die behoort tot een cysteïne en één die behoort tot de lange arm van een fosfopantheteïne; ACP bindt zich aan het substraat dat via de fosfopantheïne-arm in contact wordt gebracht met de andere enzymen die zo hun enzymatische werking kunnen uitoefenen.
Acetyl-co-enzym A (door middel van ACP-acetyltransacylase) bindt zich aan het ACP-enzym (meer precies aan de zwavel van cysteïne die het cysteylderivaat vormt) en co-enzym A komt vrij; ACP-malonyltransacylase grijpt dan in wat de aanval van malonyl op phosphopanthetheine (ook in dit proces komt co-enzym A vrij dat aanvankelijk aan malonyl gebonden was).
De volgende stap omvat het β-keto-acyl ACP-synthase dat een condenserend enzym is: het maakt de fusie tussen de twee skeletten mogelijk; malonyl wordt gemakkelijk gedecarboxyleerd en er ontstaat een carbonyl van het acetylderivaat cysteïne: het cysteïne komt vrij en er wordt een β-keto (acetylacetyl)fosfopantethinederivaat gevormd.
Vervolgens grijpt het β-keto-acyl-ACP-reductase in dat de carbonyl verder reduceert tot het ACP-enzym (een hydroxide wordt gevormd door NADPH dat wordt gereduceerd tot NADP+).
Nu werkt 3-hydroxy-acyl ACP-dehydratase (uitdroging treedt op) wat leidt tot de vorming van een onverzadigd systeem (alkeen).
Het volgende proces is het enoyl-ACP-reductase (het voert een hydrogenering uit: het alkaan wordt gevormd en NADPH wordt gereduceerd tot NADP+).
De laatste fase omvat de omzetting van het acylproduct verkregen uit de eerste cyclus in een verbinding die in staat is om een tweede cyclus te starten: het transacylase-enzym brengt de acyl over op de cysteïne, waardoor de plaats van de pantethine vrij blijft, die nu bereid zal zijn een andere te binden. malonyl.
Bij β-oxidatie wordt een molecuul FAD gebruikt om de onverzadigde α-β-metaboliet trans-enoyl-co-enzym A te verkrijgen door dehydrogenering; bij de synthese van vetzuren wordt in plaats daarvan een molecuul NADPH gebruikt om de tegenovergestelde reactie te laten plaatsvinden.
Gewoonlijk worden vetzuren met zestien koolstofatomen gesynthetiseerd, maar vetzuren met achttien, twintig of tweeëntwintig koolstofatomen kunnen ook worden geproduceerd; de vetzuren worden vervolgens veresterd om triglyceriden te vormen met geactiveerde glycerol (dwz glycerol 3-fosfaat). Dit laatste kan worden verkregen uit dihydroxyacetonfosfaat door de werking van het enzym glycerolfosfaatdehydrogenase of uit glycerol via het enzym glycerolkinase.
De gesynthetiseerde vetzuren moeten naar het vetweefsel worden gestuurd; ze worden in de bloedbaan getransporteerd in de vorm van triglyceriden of, gedeeltelijk als zodanig, met behulp van een transporteiwit dat albumine is.