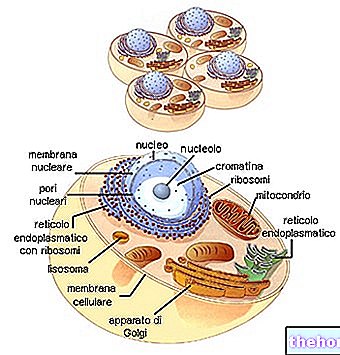
De typische structuur van het celmembraan bestaat uit een fosfolipide dubbellaag tussen twee eiwitlagen die zich ter hoogte van de scheidingsvlakken tussen de interne en externe fasen van de cel bevinden. De lipidelaag is bimoleculair, met de polaire groepen tegenover de eiwitlaag, terwijl de apolaire groepen tegenover elkaar staan met een isolatiefunctie.
Celmembranen, met een dikte van slechts 90 A, zijn niet zichtbaar onder de doorvallende lichtmicroscoop. Voor de komst van de elektronenmicroscopie gingen cytologen ervan uit dat de cel omgeven was door een onzichtbare film, want als deze hypothetische film zou worden verbroken, zou de celinhoud eruit kunnen lekken. Tegenwoordig kan met de elektronenmicroscoop het membraan worden gevisualiseerd als een dun dubbele ononderbroken lijn Volgens de huidige hypothese bestaat het membraan in wezen uit: moleculen van fosfolipiden en cholesterol zo gerangschikt dat hun hydrofobe staarten naar binnen zijn gekeerd.
De polypeptideketens van de membraaneiwitmoleculen staan loodrecht op de lipidemoleculen en er wordt aangenomen dat ze de cohesie tussen de verschillende delen van het plasmamembraan behouden.
De vliezige structuur vervult de taak om de cellulaire omgeving te scheiden van de extracellulaire, de kern van het cytoplasma, en ook het materiaal in de verschillende organellen van de cytoplasmatische matrix.
In elke cel, of deze nu dierlijk of plantaardig is, heeft de perifere laag van het protoplasma de morfologische en functionele kenmerken van een membraan dat is geplaatst om twee verschillende omgevingen te scheiden, die kunnen worden geïdentificeerd met oplossingen met verschillende chemisch-fysische kenmerken en samenstellingen. De functie van dit diafragma is om de doorgang van water en andere kleine opgeloste stoffen in de cel mogelijk te maken, terwijl het zich verzet tegen opgeloste stoffen met een hoog molecuulgewicht. In het algemeen wordt de stroomrichting bepaald door de concentratie van de samenstelling van de oplossing aan de zijkanten van het membraan, vindt de stroom altijd plaats in de richting van de meest verdunde oplossing naar de meest geconcentreerde: dat wil zeggen, het heeft de neiging om de twee concentraties in evenwicht te brengen en stopt wanneer gelijkheid is bereikt. De druk die nodig is om deze beweging volledig te stoppen, wordt osmotische druk genoemd. Het is des te groter naarmate de oplossing geconcentreerder is.
Het celmembraan is geen ideaal semi-permeabel membraan, omdat het ondoordringbaar is voor sommige, maar niet alle, aanwezige opgeloste stoffen. De permeabiliteit van het membraan voor opgeloste stoffen hangt niet uitsluitend af van zijn chemisch-fysische structurele kenmerken, maar grotendeels van verschijnselen die nauw verbonden zijn met het cellulaire metabolisme.
De cellen worden, in relatie tot hun gedrag ten opzichte van osmotische druk en omgevingsdruk, onderverdeeld in: poikilosmotisch en homoosmotisch. De eerstgenoemden hebben een osmotische druk die gelijk is aan of bijna gelijk is aan die van hun omgeving, de laatstgenoemden zijn in staat om een osmotische druk te handhaven binnen een groot aantal waarden, die sterk verschillen van de omgevingswaarden. Rekening houdend met deze kenmerken van het gedrag van dierlijke en plantaardige cellen, creëerde J. Traube een speciaal apparaat, precies bestaande uit een semipermeabel membraan, dat het gedrag van levende cellen kunstmatig moest reproduceren in het licht van gegeven oplossingen. Aanvankelijk werd een koperferrocyanidefilm als membraan gebruikt; vervolgens werden semipermeabele membranen ingebracht waarmee het mogelijk was om de entiteit van aanzienlijke osmotische drukken vast te stellen.
Tot slot kan worden gesteld dat de passage van de verschillende stoffen door het plasmamembraan kan plaatsvinden door eenvoudige diffusie, gefaciliteerd of door actief transport.
Eenvoudige diffusie: passief transport door de lipide dubbellaag. Diffusie is de beweging van moleculen van de ene zone naar de andere na hun willekeurige thermische agitatie. Bij eenvoudige diffusie wordt de permeabiliteit van het membraan bepaald door de volgende factoren: (a) de vetoplosbaarheid van de diffunderende stof, (b) de grootte en vorm van de diffunderende moleculen, (c) de temperatuur en (d) de dikte van het membraan.
Gefaciliteerde diffusie: passief transport door membraaneiwitten. Gefaciliteerde diffusie wordt aangedreven door twee soorten transporteiwitten: (a) transporters, die moleculen aan de ene kant van het membraan binden en ze naar de andere transporteren dankzij een conformationele modificatie, en (b) kanalen, die poriën vormen die zich uitstrekken van de ene kant van het membraan. kant van het membraan naar de andere. Bij gefaciliteerde diffusie wordt de permeabiliteit van het membraan bepaald door twee factoren: (a) de transportsnelheid van de afzonderlijke dragers of kanalen en (b) het aantal dragers of kanalen dat in het membraan aanwezig is.
Actief transport. Er zijn twee hoofdtypen actief transport: primair actief transport, dat gebruik maakt van ATP of andere vormen van chemische energie, en secundair actief transport, dat de elektrochemische gradiënt van een stof gebruikt als energiebron om het actieve transport van een hoge stof te induceren.

Klik op de namen van de verschillende organellen om de diepgaande studie te lezen
Afbeelding afkomstig van www.progettogea.com